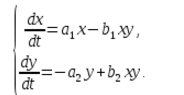
Трептения на системата хищник-жертва (модел на Лотка-Волтер). Курсова работа: Качествено изследване на модела хищник-плячка Ситуационен модел хищник-плячка
Често представители на един вид (популация) се хранят с представители на друг вид.
Моделът на Лотка-Волтера е модел на взаимното съществуване на две популации от типа "хищник-плячка".
Моделът "хищник-плячка" е получен за първи път от А. Лотка през 1925 г., който го използва, за да опише динамиката на взаимодействащите си биологични популации. През 1926 г., независимо от Лотка, подобни (при това по-сложни) модели са разработени от италианския математик В. Волтера, чиито задълбочени изследвания в областта на екологичните проблеми полагат основите на математическата теория на биологичните общности или т.нар. . математическа екология.
В математическа форма предложената система от уравнения има формата:
където x е броят на плячката, y е броят на хищниците, t е времето, α, β, γ, δ са коефициенти, които отразяват взаимодействията между популациите.
Формулиране на проблема
Да разгледаме затворено пространство, в което има две популации - тревопасни ("жертви") и хищници. Смята се, че животни не се внасят или изнасят и че има достатъчно храна за тревопасните. Тогава уравнението за промяна на броя на жертвите (само жертвите) ще приеме формата:
където $α$ е раждаемостта на жертвите,
$x$ е размерът на популацията плячка,
$\frac(dx)(dt)$ е темпът на растеж на популацията на плячката.
Когато хищниците не ловуват, те могат да измрат, така че уравнението за броя на хищниците (само хищници) става:
Където $γ$ е коефициентът на изчезване на хищници,
$y$ е размерът на популацията на хищниците,
$\frac(dy)(dt)$ е темпът на растеж на популацията на хищниците.
Когато хищници и плячка се срещнат (честотата на срещите е правопропорционална на продукта), хищниците унищожават плячка с коефициент, добре хранените хищници могат да възпроизвеждат потомство с коефициент. Така системата от уравнения на модела ще приеме формата:
Решението на проблема
Нека изградим математически модел на съвместното съществуване на две биологични популации от типа "хищник-жертва".
Нека две биологични популации живеят заедно в изолирана среда. Средата е стационарна и осигурява неограничено количество от всичко необходимо за живот на един от видовете – жертвата. Друг вид - хищник - също живее в стационарни условия, но се храни само с плячка. Котки, вълци, щуки, лисици могат да действат като хищници, а пилета, зайци, караси, мишки, съответно, могат да действат като жертви.
За категоричност нека считаме котките за хищници, а пилетата за жертви.
И така, пилетата и котките живеят в някакво изолирано пространство - домашния двор. Околната среда осигурява на пилетата неограничена храна, докато котките ядат само пилетата. Означаваме с
$x$ - броят на пилетата,
$y$ е броят на котките.
С течение на времето броят на пилетата и котките се променя, но ние ще считаме $x$ и $y$ за непрекъснати функции на времето t. Нека наречем двойка числа $x, y)$ състоянието на модела.
Нека намерим как се променя състоянието на модела $(x, y).$
Помислете за $\frac(dx)(dt)$, скоростта на промяна в броя на пилетата.
Ако няма котки, тогава броят на пилетата се увеличава и колкото по-бързо, толкова повече пилета. Ще считаме зависимостта линейна:
$\frac(dx)(dt) a_1 x$,
$a_1$ е коефициент, който зависи само от условията на живот на пилетата, тяхната естествена смъртност и раждаемост.
$\frac(dy)(dt)$ – скоростта на промяна в броя на котките (ако няма пилета), зависи от броя на котките y.
Ако няма пилета, тогава броят на котките намалява (нямат храна) и те измират. Ще считаме зависимостта линейна:
$\frac(dy)(dt) - a_2 y$.
В екосистемата скоростта на изменение на броя на всеки вид също ще се счита за пропорционална на неговата численост, но само с коефициент, зависещ от броя на индивидите от друг вид. И така, за пилетата този коефициент намалява с увеличаване на броя на котките, а за котките се увеличава с увеличаване на броя на пилетата. Ще приемем, че зависимостта също е линейна. Тогава получаваме система от диференциални уравнения:
Тази система от уравнения се нарича модел на Волтера-Лотка.
a1, a2, b1, b2 са числени коефициенти, които се наричат параметри на модела.
Както можете да видите, естеството на промяната в състоянието на модела (x, y) се определя от стойностите на параметрите. Чрез промяна на тези параметри и решаване на системата от уравнения на модела е възможно да се изследват моделите на промени в състоянието на екологичната система.
С помощта на програмата MATLAB системата от уравнения на Лотка-Волтера се решава, както следва:
На фиг. 1 е показано решението на системата. В зависимост от началните условия решенията са различни, което съответства на различни цветове на траекториите.
На фиг. 2 показва същите решения, но като се вземе предвид времевата ос t (т.е. има времева зависимост).
КОМПЮТЪРЕН МОДЕЛ "ХИЩНИК-Плячка"
Казачков Игор Алексеевич 1 , Гусева Елена Николаевна 2
1 Магнитогорски държавен технически университет на името на V.I. Г.И. Носова, Институт по строителство, архитектура и изкуство, студент 5-та година
b Магнитогорски държавен технически университет Г.И. Носова, Институт по енергетика и автоматизирани системи, кандидат на педагогическите науки, доцент на катедра "Бизнес информатика и информационни технологии"
анотация
Тази статия е посветена на прегледа на компютърния модел "хищник-плячка". Проучването ни позволява да заявим, че екологичното моделиране играе огромна роля в изучаването на околната среда. Този въпрос е многостранен.
КОМПЮТЪРЕН МОДЕЛ "ХИЩНИК-ЖЕРТВА"
Казачков Игор Алексеевич 1 , Гусева Елена Николаевна 2
1 Носов Магнитогорски държавен технически университет, Институт по строителство, архитектура и изкуства, студент от 5-ти курс
2 Магнитогорски държавен технически университет "Носов", Институт по енергетика и автоматизирани системи, доктор на педагогическите науки, доцент на катедра "Бизнес компютърни науки и информационни технологии"
Резюме
Тази статия предоставя преглед на компютърния модел "хищник-жертва". Проучването предполага, че симулацията на околната среда играе огромна роля в изследването на околната среда. Този проблем е многостранен.
Екологичното моделиране се използва за изследване на околната среда около нас. Математическите модели се използват в случаите, когато няма естествена среда и природни обекти, те помагат да се предвиди влиянието на различни фактори върху обекта, който се изследва. Този метод поема функциите на проверка, конструиране и интерпретиране на резултатите. Въз основа на такива форми екологичното моделиране се занимава с оценка на промените в околната среда около нас.
В момента такива форми се използват за изучаване на околната среда около нас, а когато е необходимо да се изследва някоя от нейните области, тогава се използва математическо моделиране. Този модел позволява да се предвиди влиянието на определени фактори върху обекта на изследване. По едно време типът „хищник-плячка“ беше предложен от учени като: Т. Малтус (Malthus 1798, Malthus 1905), Верхулст (Verhulst 1838), Пърл (Pearl 1927, 1930), както и А. Лотка ( Lotka 1925, 1927 ) и V. Volterra (Volterra 1926).Тези модели възпроизвеждат периодичния осцилационен режим, възникващ в резултат на междувидовите взаимодействия в природата.
Един от основните методи на познание е моделирането. Освен че може да предскаже промените в околната среда, той също помага да се намери най-добрият начин за решаване на проблем. Дълго време математическите модели се използват в екологията, за да установят закономерностите, тенденциите в развитието на популациите и да помогнат да се подчертае същността на наблюденията. Оформлението може да служи като пример поведение, обект.
При пресъздаване на обекти в математическата биология се използват прогнози на различни системи, предвидени са специални индивидуалности на биосистемите: вътрешната структура на индивида, условията за поддържане на живота, постоянството на екологичните системи, благодарение на което се запазва жизнената активност на системите.
Появата на компютърната симулация значително разшири границата на изследователските възможности. Имаше възможност за многостранно прилагане на трудни форми, които не позволяват аналитично изследване, появиха се нови тенденции, както и симулационно моделиране.
Нека разгледаме какво е обектът на моделиране. „Обектът е затворено местообитание, където се осъществява взаимодействието на две биологични популации: хищници и плячка. Протича процесът на растеж, изчезване и размножаванедиректно върху повърхността на околната среда. Плячката се храни с ресурсите, които присъстват в околната среда, докато хищниците се хранят с плячка. В същото време хранителните ресурси могат да бъдат както възобновяеми, така и невъзобновяеми.
През 1931 г. Вито Волтера извежда следните закони на връзката хищник-плячка.
Законът на периодичния цикъл - процесът на унищожаване на плячката от хищник често води до периодични колебания в броя на популациите и на двата вида, в зависимост само от скоростта на растеж на месоядните и тревопасните животни и от първоначалното съотношение на техния брой .
Закон за запазване на средните стойности - средното изобилие на всеки вид е постоянно, независимо от първоначалното ниво, при условие че специфичните темпове на нарастване на популацията, както и ефективността на хищничеството, са постоянни.
Законът за нарушаване на средните стойности - с намаляване на двата вида пропорционално на техния брой, средната популация на плячката се увеличава, а хищниците - намалява.
Моделът хищник-плячка е специална връзка между хищника и плячката, в резултат на което и двамата печелят. Оцеляват най-здравите и адаптирани към условията на околната среда индивиди, т.е. Всичко това се дължи на естествения подбор. В среда, в която няма възможност за възпроизводство, хищникът рано или късно ще унищожи популацията на плячката, след което самата тя ще измре.
На земята има много живи организми, които при благоприятни условия увеличават броя на роднините си до огромни размери. Тази способност се нарича: биотичен потенциал на вида, т.е. увеличаване на популацията на даден вид за даден период от време. Всеки вид има свой собствен биотичен потенциал, например големите видове организми могат да растат само 1,1 пъти годишно, докато организмите от по-малки видове, като ракообразни и др. могат да увеличат появата си до 1030 пъти, но бактериите са още по-големи. Във всеки от тези случаи населението ще нараства експоненциално.
Експоненциалният растеж на населението е геометрична прогресия на растежа на населението. Тази способност може да се наблюдава в лабораторията при бактерии, дрожди. В извънлабораторни условия може да се наблюдава експоненциален растеж при скакалци или други видове насекоми. Такова увеличение на числеността на вида може да се наблюдава в онези места, където практически няма врагове и има повече от достатъчно храна. В крайна сметка растежът на вида, след като популацията се увеличи за кратко време, растежът на популацията започна да намалява.
Помислете за компютърен модел на възпроизвеждане на бозайници на примера на модела Lotka-Volterra. Позволявам два вида животни живеят в определен район: елени и вълци. Математически модел на промяната на населението в моделаТрайс-Волтера:
Първоначалният брой на жертвите е xn, броят на хищниците е yn.
Параметри на модела:
P1 е вероятността за среща с хищник,
P2 е темпът на растеж на хищниците за сметка на плячката,
d е коефициентът на смъртност на хищниците,
а е нарастването на броя на жертвите.
В задачата за обучение бяха зададени следните стойности: броят на елените беше 500, броят на вълците беше 10, скоростта на растеж на елените беше 0,02, скоростта на растеж на вълците беше 0,1, вероятността за среща с хищник беше 0,0026, скоростта на растеж на хищниците поради плячка е 0,000056. Данните са изчислени за 203 години.
Изследване на влиянието скоростта на нарастване на жертвите за развитието на две популации, останалите параметри ще бъдат оставени непроменени.В схема 1 се наблюдава увеличаване на броя на плячката, а след това с известно забавяне се наблюдава увеличаване на хищниците. След това хищниците избиват плячката, броят на плячката рязко намалява, последвано от намаляване на броя на хищниците (фиг. 1).

Фигура 1. Размер на населението с ниска раждаемост сред жертвите
Нека анализираме промяната в модела чрез увеличаване на раждаемостта на жертвата a=0,06. В схема 2 виждаме цикличен осцилационен процес, водещ до увеличаване на броя на двете популации с течение на времето (фиг. 2).

Фигура 2. Размер на населението при средна раждаемост на жертвите
Нека разгледаме как ще се промени динамиката на популациите при висока стойност на раждаемостта на жертвата a = 1,13. На фиг. 3, има рязко увеличение на числеността и на двете популации, последвано от изчезване както на плячка, така и на хищник. Това се дължи на факта, че популацията на жертвите се е увеличила до такава степен, че ресурсите са започнали да се изчерпват, в резултат на което жертвата умира. Изчезването на хищниците се дължи на факта, че броят на жертвите е намалял и хищниците са изчерпали ресурсите си за съществуване.

Фигура 3. Популации с висока раждаемост в плячка
Въз основа на анализа на данните от компютърни експерименти можем да заключим, че компютърното моделиране ни позволява да прогнозираме размера на популациите, да изследваме влиянието на различни фактори върху динамиката на популацията. В горния пример изследвахме модела хищник-плячка, ефекта от раждаемостта на плячката върху броя на елените и вълците. Малкото увеличение на популацията на плячката води до малко увеличение на плячката, която след определен период се унищожава от хищници.Умереното увеличение на популацията на плячката води до увеличаване на размера и на двете популации. Голямото увеличение на популацията на плячка първо води до бързо нарастване на популацията на плячка, това се отразява на увеличаването на растежа на хищниците, но след това размножаващите се хищници бързо унищожават популацията на елените. В резултат на това и двата вида изчезват.
Още през 20-те години. A. Lotka и малко по-късно, независимо от него, V. Volterra предложи математически модели, описващи съчетани колебания в размера на популациите на хищници и жертви. Помислете за най-простата версия на модела Lotka-Volterra. Моделът се основава на редица предположения:
1) популацията на плячката в отсъствието на хищник нараства експоненциално,
2) натискът на хищниците възпрепятства този растеж,
3) смъртността на плячката е пропорционална на честотата на срещи между хищник и плячка (или по друг начин, пропорционална на произведението на гъстотата на популацията им);
4) раждаемостта на хищник зависи от интензивността на консумация на плячка.
Моментната скорост на промяна в популацията на плячката може да бъде изразена чрез уравнението
dN ямка /dt = r 1 N ямка - p 1 N ямка N x,
където r 1 - специфична моментна скорост на нарастване на популацията на плячката, p 1 - константа, свързваща смъртността на плячката с гъстотата на хищника, a н и N x - плътност съответно на плячка и хищник.
Моментният темп на растеж на популацията на хищници в този модел се приема, че е равен на разликата между раждаемостта и постоянната смъртност:
dN x / dt \u003d p 2 N f N x - d 2 N x,
където p2 - константа, свързваща раждаемостта в популацията на хищниците с плътността на плячката, a d 2 - специфична смъртност на хищник.
Съгласно горните уравнения всяка от взаимодействащите популации в нарастването си е ограничена само от другата популация, т.е. увеличаването на броя на плячката е ограничено от натиска на хищниците, а увеличаването на броя на хищниците е ограничено от недостатъчния брой плячка. Не се предполагат самоограничаващи се популации. Смята се например, че винаги има достатъчно храна за жертвата. Също така не се очаква популацията на плячка да излезе извън контрола на хищника, въпреки че всъщност това се случва доста често.
Въпреки цялата условност на модела на Лотка-Волтера, той заслужава внимание дори само защото показва как дори такава идеализирана система на взаимодействие между две популации може да генерира доста сложна динамика на техния брой. Решението на системата от тези уравнения ни позволява да формулираме условията за поддържане на постоянно (равновесно) изобилие на всеки от видовете. Популацията на плячката остава постоянна, ако плътността на хищниците е r 1 /p 1, а за да остане популацията на хищниците постоянна, плътността на плячката трябва да е равна на d 2 /p 2 . Ако на графиката нанесем плътността на жертвите по абсцисата н и , а по оста y - плътността на хищника н х, тогава изоклините, показващи състоянието на постоянството на хищника и плячката, ще бъдат две прави линии, перпендикулярни една на друга и на координатните оси (фиг. 6а). Предполага се, че под определена плътност на плячката (равна на d 2 /p 2) плътността на хищника винаги ще намалява, а над нея винаги ще нараства. Съответно, плътността на плячката се увеличава, ако плътността на хищника е под стойността, равна на r 1 /p 1, и намалява, ако е над тази стойност. Пресечната точка на изоклините съответства на условието за постоянство на броя на хищниците и плячката, а други точки на равнината на тази графика се движат по затворени траектории, като по този начин отразяват регулярните колебания в броя на хищниците и плячката (фиг. 6, б).Диапазонът на колебанията се определя от първоначалното съотношение на плътността на хищника и плячката. Колкото по-близо е до пресечната точка на изоклините, толкова по-малък е кръгът, описан от векторите, и съответно по-малка е амплитудата на трептенията.
Ориз. 6. Графично представяне на модела на Лотка-Волтер за системата хищник-жертва.
Един от първите опити за получаване на колебания в броя на хищниците и плячката в лабораторни експерименти принадлежи на G.F. Гаузе. Обектите на тези експерименти са парамециеви реснички (парамеций caudatum) и хищните реснички didinium (дидиний nasutum). Суспензия от бактерии, редовно въвеждана в средата, служи като храна за парамеция, докато дидиниумът се храни само с парамеция. Тази система се оказа изключително нестабилна: натискът на хищника, тъй като броят му се увеличи, доведе до пълното унищожаване на жертвите, след което популацията на самия хищник също измря. Усложнявайки експериментите, Гауз организира подслон за жертвата, като въвежда малко стъклена вата в епруветките с ресничките. Сред нишките на памучната вата парамецията можеше да се движи свободно, но дидиниумът не можеше. В тази версия на експеримента дидиниумът изяжда целия парамеций, плаващ в частта на епруветката, свободна от памучна вата, и измира, а популацията на парамеция след това се възстановява поради възпроизвеждането на индивиди, оцелели в приюта. Гауз успява да постигне известно подобие на колебания в броя на хищниците и плячката само когато от време на време въвежда и плячка, и хищник в културата, като по този начин симулира имиграция.
40 години след работата на Гаузе, неговите експерименти бяха повторени от Л. Лакинбиял (Лъкинбил), който използва реснички като жертва парамеций аурелия, но като хищник на същ дидиний nasutum. Luckinbill успя да получи няколко цикъла на колебания в изобилието на тези популации, но само в случай, че плътността на парамецията е ограничена от липса на храна (бактерии) и към културалната течност е добавена метилцелулоза, вещество, което намалява скорост както на хищника, така и на жертвата и следователно намалява тяхната честота.възможни срещи. Оказа се също, че е по-лесно да се постигнат колебания между хищника и плячката, ако се увеличи обемът на експерименталния съд, въпреки че условието за ограничаване на храната на плячката също е необходимо в този случай. Ако обаче към системата от хищник и плячка, съжителстващи в колебателен режим, се добави излишна храна, тогава отговорът беше бързо увеличаване на броя на плячката, последвано от увеличаване на броя на хищника, което от своя страна води до пълното унищожаване на популацията на плячката.
Моделите на Лотка и Волтера дадоха тласък на развитието на редица други по-реалистични модели на системата хищник-плячка. По-специално, доста прост графичен модел, който анализира съотношението на различни изоклини на плячка хищник, е предложен от М. Розенцвайг и Р. Макартър (Rosenzweig, MacArthur). Според тези автори стационарният ( = постоянно) изобилието на плячката в координатните оси на хищника и плътността на плячката може да бъде представено като изпъкнала изоклина (фиг. 7а). Едната точка на пресичане на изоклината с оста на плътността на плячката съответства на минималната допустима плътност на плячката (ниската популация е изложена на много висок риск от изчезване, макар и само поради ниската честота на срещи между мъжки и женски), а другата е максимумът, определен от количеството налична храна или поведенческите характеристики на самата плячка. Подчертаваме, че все пак говорим за минимална и максимална плътност при липса на хищник. Когато се появи хищник и числеността му се увеличи, минималната допустима плътност на плячката очевидно трябва да бъде по-висока, а максималната - по-ниска. Всяка стойност на плътността на плячката трябва да съответства на определена плътност на хищника, при която популацията на плячката е постоянна. Географското място на такива точки е изоклина на плячката в координатите на плътността на хищника и плячката. Векторите, показващи посоката на промените в плътността на плячката (ориентирани хоризонтално), имат различни посоки от различните страни на изоклина (фиг. 7а).

Ориз. Фиг. 7. Изоклини на стационарни популации на плячка (а) и хищник (б).
За хищника е построена и изоклина в същите координати, съответстваща на стационарното състояние на популацията му. Векторите, показващи посоката на промяна в изобилието на хищници, са ориентирани нагоре или надолу в зависимост от това от коя страна на изоклина се намират. Изоклинната форма на хищника, показана на фиг. 7, b.се определя, първо, от наличието на определена минимална плътност на плячката, достатъчна за поддържане на популацията на хищника (при по-ниска плътност на плячката, хищникът не може да увеличи изобилието си), и второ, от наличието на определена максимална плътност на самия хищник , над което изобилието ще намалее независимо от изобилието на жертвите.

Ориз. 8. Възникване на колебателни режими в системата хищник-жертва в зависимост от разположението на изоклините на хищник и жертва.
При комбиниране на изоклините на плячката и хищника на една графика са възможни три различни варианта (фиг. 8). Ако изоклината на хищника пресича изоклина на плячката в точката, където тя вече намалява (при висока плътност на плячка), векторите, показващи промяната в изобилието на хищник и плячка, образуват траектория, която се извива навътре, което съответства на затихнали флуктуации в изобилието от плячка и хищник (фиг. 8, а). В случай, когато изоклината на хищника пресича изоклина на плячката в нейната възходяща част (т.е. в областта на ниски стойности на плътност на плячката), векторите образуват развиваща се траектория и възникват съответно колебания в изобилието на хищник и плячка, с нарастваща амплитуда (фиг. 8, б).Ако изоклина на хищник пресича изоклина на плячка в областта на нейния връх, тогава векторите образуват порочен кръг, а колебанията в броя на плячката и хищника се характеризират със стабилна амплитуда и период (фиг. 8, в).
С други думи, затихналите колебания съответстват на ситуация, в която хищник значително засяга популация на плячка, която е достигнала само много висока плътност (близо до границата), докато колебания с нарастваща амплитуда възникват, когато хищникът е в състояние бързо да увеличи числеността си дори при ниска плътност на плячката и такива я унищожават бързо. В други версии на своя модел Позенцвайг и Макартър показаха, че трептенията хищник-плячка могат да бъдат стабилизирани чрез въвеждане на „убежище“, т.е. ако приемем, че в зона с ниска плътност на плячка има зона, в която броят на плячката нараства независимо от броя на присъстващите хищници.
Желанието да се направят моделите по-реалистични, като се направят по-сложни, се проявява в трудовете не само на теоретиците, но и експериментатори. По-специално, интересни резултати бяха получени от Huffaker, който показа възможността за съвместно съществуване на хищник и плячка в колебателен режим, използвайки примера на малък тревопасен кърлеж. Еотетранихус sexmaculatusи хищен кърлеж, който го напада Тифлодромус occidentalis. Като храна за тревопасните акари са използвани портокали, поставени върху тави с дупки (като тези, използвани за съхранение и транспортиране на яйца). В оригиналната версия имаше 40 дупки на една тава, като някои от тях съдържаха портокали (частично обелени), а други с гумени топки. И двата вида кърлежи се размножават партеногенетично много бързо и следователно естеството на динамиката на тяхната популация може да бъде разкрито за относително кратък период от време. След като постави 20 женски от тревопасния кърлеж върху табла, Huffaker наблюдава бързо нарастване на популацията му, която се стабилизира на ниво от 5-8 хиляди индивида (на един портокал). Ако няколко индивида от хищник бяха добавени към нарастващата популация на плячка, тогава популацията на последната бързо увеличи броя си и измря, когато всички жертви бяха изядени.
Чрез увеличаване на размера на тавата до 120 дупки, в които отделни портокали са произволно разпръснати сред много гумени топки, Huffaker успя да разшири съвместното съществуване на хищник и плячка. Важна роля във взаимодействието между хищник и плячка, както се оказа, играе съотношението на скоростта на тяхното разпръскване. Huffaker предположи, че чрез улесняване на движението на плячката и затрудняване на движението на хищника е възможно да се увеличи времето на съвместното им съществуване. За да направите това, 6 портокала бяха произволно поставени върху тава със 120 дупки сред гумени топки и вазелинови бариери бяха поставени около дупките с портокали, за да се предотврати настаняването на хищника, а за да се улесни настаняването на жертвата, бяха укрепени дървени колчета таблата, която служи като вид "платформи за излитане" за тревопасни акари (факт е, че този вид освобождава тънки нишки и с тяхна помощ може да се издига във въздуха, разпространявайки се във вятъра). В такова сложно местообитание хищник и плячка съжителстват в продължение на 8 месеца, демонстрирайки три пълни цикъла на колебания в изобилието. Най-важните условия за това съвместно съществуване са следните: хетерогенност на местообитанието (в смисъл на наличие на зони, подходящи и неподходящи за плячката в него), както и възможността за миграция на плячка и хищник (при запазване на известно предимство на плячката в скоростта на този процес). С други думи, хищникът може напълно да унищожи едно или друго местно натрупване на плячка, но някои от жертвите ще имат време да мигрират и да създадат други местни натрупвания. Рано или късно хищникът ще стигне и до нови местни клъстери, но междувременно плячката ще има време да се установи на други места (включително тези, където е живял преди, но след това е бил унищожен).
Нещо подобно на това, което Huffaker наблюдава в експеримента, се случва и в естествени условия. Така например пеперуда от кактусов молец (Cactoblastis cactorum), донесен в Австралия, значително намали броя на кактуса опунция, но не го унищожи напълно именно защото кактусът успява да се установи малко по-бързо. На местата, където опунцията е напълно унищожена, огненият молец също престава да се среща. Следователно, когато след известно време бодливата круша отново проникне тук, тогава за определен период тя може да расте без риск да бъде унищожена от молец. С течение на времето обаче молецът отново се появява тук и, бързо размножавайки се, унищожава бодливата круша.
Говорейки за колебанията между хищници и жертви, не можем да не споменем цикличните промени в числеността на зайците и рисовете в Канада, проследени от статистиката за добива на кожи от Hudson Bay Company от края на 18 век до началото на 20 век. век. Този пример често се разглежда като класическа илюстрация на флуктуациите хищник-плячка, въпреки че всъщност виждаме само растеж на популацията на хищник (рис) след растежа на плячка (заек). Що се отнася до намаляването на броя на зайците след всяко издигане, то не може да се обясни само с повишения натиск на хищниците, а е свързано с други фактори, очевидно най-вече с липсата на храна през зимата. До този извод стигна по-специално М. Гилпин, който се опита да провери дали тези данни могат да бъдат описани с класическия модел на Лотка-Волтера. Резултатите от теста показаха, че няма задоволително напасване на модела, но колкото и да е странно, ставаше по-добре, ако хищникът и плячката бяха разменени, т.е. рисът се тълкува като "жертва", а заекът - като "хищник". Подобна ситуация беше отразена в закачливото заглавие на статията („Ядат ли зайците рисове?“), която по същество е много сериозна и публикувана в сериозно научно списание.
Модел на ситуация тип "хищник-жертва".
Нека разгледаме математически модел на динамиката на съвместното съществуване на два биологични вида (популации), взаимодействащи помежду си според типа „хищник-плячка“ (вълци и зайци, щуки и каракуди и др.), наречен Волтер- Модел Lotka. За първи път е получен от А. Лотка (1925 г.), а малко по-късно и независимо от Лотка подобни и по-сложни модели са разработени от италианския математик В. Волтера (1926 г.), чиято работа всъщност полага основите на т.н. наречена математическа екология.
Да предположим, че има два биологични вида, които живеят заедно в изолирана среда. Това предполага:
- 1. Жертвата може да намери достатъчно храна, за да живее;
- 2. При всяка среща на жертвата с хищника, последният убива жертвата.
За категоричност ще ги наречем караси и щуки. Позволявам
състоянието на системата се определя от количествата x(t)и y(t)- броят на караси и щуки в момента Ж.За да получим математически уравнения, които приблизително описват динамиката (промяната във времето) на популацията, процедираме по следния начин.
Както в предишния модел на нарастване на населението (вижте раздел 1.1), за жертвите имаме уравнението
където а> 0 (раждаемостта надвишава смъртността)
Коефициент аувеличаването на плячката зависи от броя на хищниците (намалява с увеличаването им). В най-простия случай a- a - fjy (a>0, p>0).Тогава за размера на популацията плячка имаме диференциалното уравнение
За популацията от хищници имаме уравнението

където b>0 (смъртността надвишава раждаемостта).
Коефициент bИзчезването на хищници намалява, ако има плячка за хранене. В най-простия случай човек може да вземе b - y -Sx (y > 0, С> 0). Тогава за размера на популацията от хищници получаваме диференциалното уравнение

По този начин уравнения (1.5) и (1.6) представляват математически модел на разглеждания проблем за взаимодействието на популациите. В този модел променливите x,y- състоянието на системата, а коефициентите характеризират нейната структура. Нелинейната система (1.5), (1.6) е моделът на Волтер-Лотка.
Уравнения (1.5) и (1.6) трябва да бъдат допълнени с начални условия - дадени стойности на първоначалните популации.
Нека сега анализираме изградения математически модел.
Нека построим фазовия портрет на системата (1.5), (1.6) (според смисъла на задачата х> 0, v > 0). Разделяйки уравнение (1.5) на уравнение (1.6), получаваме уравнение с разделими променливи
Използвайки това уравнение, ще имаме
Съотношението (1.7) дава уравнението на фазовите траектории в неявна форма. Система (1.5), (1.6) има стационарно състояние, определено от

От уравнения (1.8) получаваме (тъй като l* Е 0, y* Е 0)
Равенствата (1.9) определят положението на равновесието на фазовата равнина (точката О)(Фигура 1.6).

От тези съображения може да се определи посоката на движение по фазовата траектория. Нека има малко шаран. g.u. x ~ 0, тогава от уравнение (1.6) y
Всички фазови траектории (с изключение на точката 0) затворени криви, обхващащи равновесното положение. Състоянието на равновесие съответства на постоянен брой x' и y' караси и щуки. Шаранът се размножава, щуката ги яде, измира, но броят на тези и другите не се променя. „Затворените фазови траектории съответстват на периодична промяна в броя на караси и щуки. Освен това траекторията, по която се движи фазовата точка, зависи от началните условия. Помислете как се променя състоянието по фазовата траектория. Нека точката е в позиция И(фиг. 1.6). Тук има малко шаран, много щука; щуките нямат какво да ядат и постепенно измират и почти
напълно изчезват. Но броят на каракудите също намалява почти до нула и
едва по-късно, когато щуката стана по-малко от при, започва увеличаването на броя на караси; скоростта им на растеж се увеличава и броят им се увеличава - това се случва приблизително до точката AT.Но увеличаването на броя на каракудите води до забавяне на процеса на изчезване на шук и техният брой започва да расте (има повече храна) - парцел слънцеОсвен това има много щуки, те ядат каракуди и ядат почти всички (раздел CD).След това щуките започват отново да измират и процесът се повтаря с период от около 5-7 години. На фиг. 1.7 качествено изградени криви на промените в броя на караси и щуки в зависимост от времето. Максимумите на кривите се редуват, като максимумите на изобилието на щуката изостават от тези на каракуда.

Това поведение е типично за различни системи хищник-плячка. Нека сега интерпретираме получените резултати.
Въпреки факта, че разглежданият модел е най-простият и в действителност всичко се случва много по-сложно, той даде възможност да се обяснят някои от мистериозните неща, които съществуват в природата. Разбираеми са разказите на риболовци за периодите, когато „щуките сами скачат в ръцете им“, обяснена е честотата на хроничните заболявания и др.
Отбелязваме още едно интересно заключение, което може да се направи от фиг. 1.6. Ако в точката Рима бърз улов на щука (в друга терминология - отстрел на вълци), след което системата "скача" до точката Q,и по-нататъшното движение се извършва по по-малка затворена траектория, което се очаква интуитивно. Ако намалим броя на щуките в точката R,тогава системата ще премине към точката С,и по-нататъшното движение ще се случи по по-голяма траектория. Амплитудата на трептене ще се увеличи. Това противоречи на интуицията, но просто обяснява такъв феномен: в резултат на отстрела на вълци броят им се увеличава с времето. Следователно изборът на момента на заснемане е важен в случая.
Да предположим, че две популации насекоми (например листна въшка и калинка, която яде листни въшки) са били в естествено равновесие. x-x*, y = y*(точка Она фиг. 1.6). Помислете за въздействието на еднократно приложение на инсектицид, което убива x> 0 от жертвите и y > 0 на хищници, без да ги унищожи напълно. Намаляването на броя на двете популации води до факта, че представляващата точка от позицията О"скача" по-близо до произхода, където x > 0, y 0 (фиг. 1.6) От това следва, че в резултат на действието на инсектицид, предназначен да унищожи плячка (листни въшки), броят на плячката (листни въшки) се увеличава, а броят на хищниците (калинките) намалява. Оказва се, че броят на хищниците може да стане толкова малък, че да изчезнат напълно по други причини (суша, болест и др.). По този начин използването на инсектициди (освен ако те почти напълно унищожават вредните насекоми) в крайна сметка води до увеличаване на популацията на тези насекоми, чийто брой е бил контролиран от други насекоми-хищници. Такива случаи са описани в книги по биология.
Като цяло темпът на нарастване на броя на жертвите азависи от L" и y: а= a(x, y) (поради наличието на хищници и хранителните ограничения).
С малка промяна в модела (1.5), (1.6), малки членове се добавят към дясната страна на уравненията (като се вземе предвид, например, конкуренцията на караси за храна и щуки за караси)
тук 0 f.i « 1.
В този случай изводът за периодичността на процеса (връщане на системата в изходно състояние), валиден за модела (1.5), (1.6), губи своята валидност. В зависимост от вида на малки корекции / и жСитуациите, показани на фиг. 1.8.

В случай (1) равновесното състояние Остабилно. За всякакви други начални условия точно тази стойност се установява след достатъчно дълго време.
В случай (2) системата "отива на пода". Стационарното състояние е нестабилно. Такава система в крайна сметка попада в такъв диапазон от стойности хи y че моделът вече не е приложим.
В случай (3) в система с неустойчиво стационарно състояние Опериодичният режим се установява с времето. За разлика от оригиналния модел (1.5), (1.6), в този модел устойчивият периодичен режим не зависи от началните условия. Първоначално малко отклонение от стабилно състояние Оводи до малки колебания О, както в модела на Volterra-Lotka, но до трептения с добре дефинирана (и независима от малката отклонение) амплитуда.
В И. Арнолд нарича модела Волтера-Лотка твърд, т.к малката му промяна може да доведе до изводи, различни от дадените по-горе. За да прецените коя от ситуациите, посочени на фиг. 1.8 е внедрена в тази система, допълнителна информация за системата е абсолютно необходима (за вида на малките корекции / и ж).
Моделът на Колмогоров прави едно важно предположение: тъй като се предполага, че това означава, че има механизми в популацията на плячката, които регулират изобилието им дори в отсъствието на хищници.
За съжаление, подобна формулировка на модела не ни позволява да отговорим на въпроса, около който напоследък има много спорове и който вече споменахме в началото на главата: как популацията на хищник може да упражнява регулаторно влияние върху жертва население, така че цялата система да е стабилна? Затова ще се върнем към модел (2.1), в който няма механизми за саморегулация (например регулиране чрез вътревидова конкуренция) в популацията на плячката (както и в популацията на хищниците); следователно единственият механизъм за регулиране на изобилието от видове в дадена общност са трофичните взаимоотношения между хищници и плячка.
Тук (така че, за разлика от предишния модел, естествено е решенията (2.1) да зависят от специфичния тип трофична функция, която от своя страна се определя от естеството на хищничеството, т.е. трофичната стратегия на хищника и отбранителната стратегия на плячката.Общи за всички тези функции (виж Фиг. I) са следните свойства:
Система (2.1) има една нетривиална стационарна точка, чиито координати се определят от уравненията
![]()
с естествено ограничение.
Има още една стационарна точка (0, 0), съответстваща на тривиалното равновесие. Лесно е да се покаже, че тази точка е седло, а координатните оси са сепаратриси.
Характеристичното уравнение за точка има формата
![]()
Очевидно за класическия модел Volterra.
Следователно стойността на f може да се разглежда като мярка за отклонението на разглеждания модел от този на Волтера.
![]()
стационарната точка е фокусът и в системата се появяват трептения; когато е изпълнено обратното неравенство, това е възел и в системата няма трептения. Устойчивостта на това равновесно състояние се определя от условието
т.е. зависи основно от вида на трофичната функция на хищника.
Условието (5.5) може да се тълкува по следния начин: за стабилността на нетривиалното равновесие на системата хищник-жертва (и следователно за съществуването на тази система) е достатъчно в близост до това състояние относителният дял на консумираната плячка от хищника се увеличава с увеличаването на броя на плячката. Наистина, делът на плячката (от общия им брой), консумирана от хищник, се описва от диференцируема функция, чието условие за растеж (позитивност на производната) изглежда като
![]()
Последното условие, взето в точката, не е нищо друго освен условието (5.5) за устойчивост на равновесие. При непрекъснатост, той също трябва да се поддържа в някакъв квартал на точката. Така, ако броят на жертвите в този квартал, тогава
Сега нека трофичната функция V има формата, показана на фиг. 11а (характеристика на безгръбначните). Може да се покаже, че за всички крайни стойности (тъй като е изпъкнал нагоре)
т.е. неравенството (5.5) не е изпълнено за никакви стойности на стационарния брой жертви.
Това означава, че в система с този тип трофична функция няма стабилно нетривиално равновесие. Възможни са няколко изхода: или броят както на плячката, така и на хищника нараства неограничено, или (когато траекторията минава близо до една от координатните оси), поради случайни причини, броят на плячката или броят на хищника ще стане равно на нула. Ако плячката умре, хищникът ще умре след известно време, но ако хищникът умре първи, тогава броят на плячката ще започне да нараства експоненциално. Третият вариант - възникването на устойчив граничен цикъл - е невъзможен, което лесно се доказва.
Наистина изразът

в положителния квадрант винаги е положителен, освен ако няма формата, показана на фиг. 11, а. Тогава, според критерия на Дюлак, в тази област няма затворени траектории и не може да съществува устойчив граничен цикъл.
И така, можем да заключим: ако трофичната функция има формата, показана на фиг. 11а, тогава хищникът не може да бъде регулатор, който осигурява стабилността на популацията на плячката и по този начин стабилността на цялата система като цяло. Системата може да бъде стабилна само ако популацията на плячката има свои собствени вътрешни регулаторни механизми, като вътревидова конкуренция или епизоотии. Тази опция за регулиране вече беше разгледана в §§ 3, 4.
По-рано беше отбелязано, че този тип трофична функция е характерна за насекомите хищници, чиито "жертви" също обикновено са насекоми. От друга страна, наблюденията върху динамиката на много природни съобщества от типа „хищник-плячка“, включително видове насекоми, показват, че те се характеризират с много големи амплитудни колебания и много специфичен тип.
Обикновено след повече или по-малко постепенно увеличаване на броя (което може да се случи или монотонно, или под формата на колебания с нарастваща амплитуда), настъпва рязък спад (фиг. 14), след което моделът се повтаря. Очевидно този характер на динамиката на изобилието от видове насекоми може да се обясни с нестабилността на тази система при ниски и средни стойности на изобилие и действието на мощни вътрешнопопулационни регулатори на изобилието при големи стойности.

Ориз. Фиг. 14. Динамика на популацията на австралийския псилид Cardiaspina albitextura, хранещ се с евкалипти. (От статията: Clark L. R. Динамиката на популацията на Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, no. 3, p. 362-380.)
Ако системата „хищник-плячка“ включва видове, способни на доста сложно поведение (например хищниците са способни да учат или плячката може да намери подслон), тогава в такава система може да съществува стабилно нетривиално равновесие. Доказателството на това твърдение е съвсем просто.
Всъщност трофичната функция трябва да има формата, показана на фиг. 11, c. Точката на тази графика е точката на контакт на правата, изтеглена от началото на координатите, с графиката на трофичната функция.Очевидно е, че в тази точка функцията има максимум. Също така е лесно да се покаже, че условието (5.5) е изпълнено за всички. Следователно, нетривиално равновесие, в което броят на жертвите е по-малък, ще бъде асимптотично стабилно
Не можем обаче да кажем нищо за това колко голяма е областта на стабилност на това равновесие. Например, ако има нестабилен граничен цикъл, тогава този регион трябва да лежи вътре в цикъла. Или друг вариант: нетривиалното равновесие (5.2) е нестабилно, но има стабилен граничен цикъл; в този случай може да се говори и за стабилност на системата хищник-жертва. Тъй като изразът (5.7) при избора на трофична функция като фиг. 11, в може да промени знака при промяна на , тогава критерият на Дюлак не работи тук и въпросът за съществуването на гранични цикли остава открит.
